
最新の研究
- 2026.06.29
- 北口研究室
抗体は「測る」から「動かす」へ
生命現象は、さまざまなタンパク質の働きによって支えられている。例えば、細胞に存在する受容体タンパク質は、外界からの刺激や周囲の環境変化を認識して、その情報を細胞内シグナルとして伝達する。こうしたシグナル伝達は、細胞の増殖、分化、代謝、細胞死、分泌など、多様な細胞機能を調節しており、発生、形態形成、恒常性維持、組織修復、神経活動といった複雑な生命現象を支える重要な仕組みである。したがって、シグナル伝達を人為的に操作できれば、どのシグナルが多様な細胞機能をどのように制御しているのかを調べることが可能となり、未解明な生命現象の理解を深めることができる。さらに、シグナル伝達の破綻に伴う細胞機能の異常を適切に回復できれば、新たな治療法の開発にもつながる。
近年、人工的に設計した受容体と、その受容体を選択的に活性化する低分子リガンドを組み合わせることで、細胞機能を操作する化学遺伝学ツールDREADDが活躍している。このツールは、特定のシグナル伝達を外部から制御できる一方、刺激として利用できるリガンドが限られている。そのため、研究者が自ら選択した分子で細胞機能を操作できるツールが望まれており、その実現には、任意の分子を認識して応答するタンパク質スイッチを設計する新たな戦略が必要であった。本研究では、抗体の優れた分子認識能を利用し、酵素の活性を抗原結合に応じて制御する酵素スイッチ「Switchbody」を開発した[1]。酵素は、さまざまな細胞内シグナルの制御に関与しており、この設計戦略を細胞内シグナル分子の産生や分解に関わる酵素へ展開できれば、任意の分子の認識によって細胞機能を制御する新たな分子ツールを実現できる。
Switchbodyの設計には、我々が研究を進めてきた蛍光免疫センサーQ-bodyの作動原理から着想を得た。Q-bodyでは、抗体に結合した蛍光色素が抗原結合部位の近くで一時的にトラップされ、蛍光が抑えられる。抗原が結合すると蛍光色素がリリースされて、蛍光が増加するため、抗原濃度を蛍光応答として定量できる。この作動原理は、抗体が単に抗原を認識するだけでなく、抗原結合に伴って、抗原結合部位の近くに配置した分子の状態を切り替えられることを示している。そこで本研究では、この作動原理を蛍光色素から酵素断片へと拡張し、抗原結合を酵素活性の変化として結びつける分子設計を試みた。
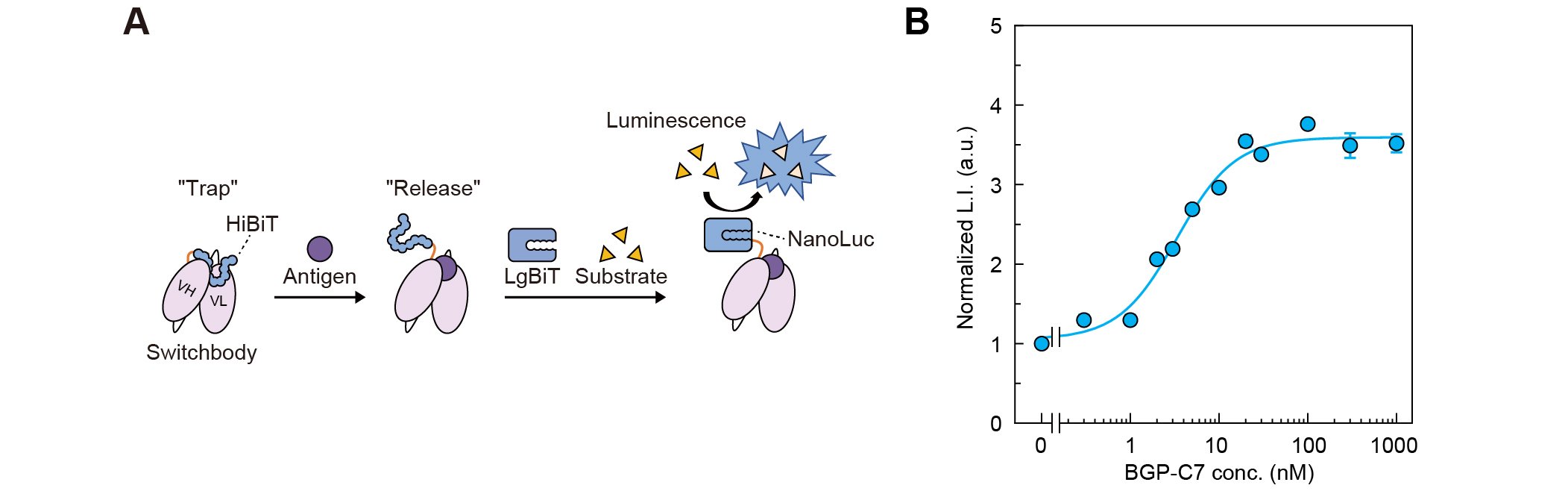
発光型Switchbodyとして、生物発光酵素NanoLucに由来する11アミノ酸の短い断片HiBiTを、ペプチド抗原(BGP-C7)を認識する抗体KTM219由来の一本鎖抗体断片scFvのN末端側に融合した分子を設計した。これは、抗原が存在しない状態では、HiBiTが抗原結合部位の近くでトラップされ、NanoLucのもう片方の断片であるLgBiTと相互作用しにくく、一方、抗原が結合するとHiBiTがリリースされ、LgBiTとの相互作用を介してNanoLucが再構成され、酵素活性を示すことを想定している(図1A)。実際に設計したHiBiT融合抗体を大腸菌で発現させ精製し、LgBiTと基質であるフリマジンの存在下で発光強度を測定したところ、抗原濃度依存的にシグナルの増大が確認され、想定どおり発光型Switchbodyとして機能することがわかった(図1B)。
 |
|
| 図1. | (A)Switchbodyの模式図(B)抗原を添加したときのSwitchbodyの発光強度変化(DOI:10.1002/advs.202508422より一部改変) |
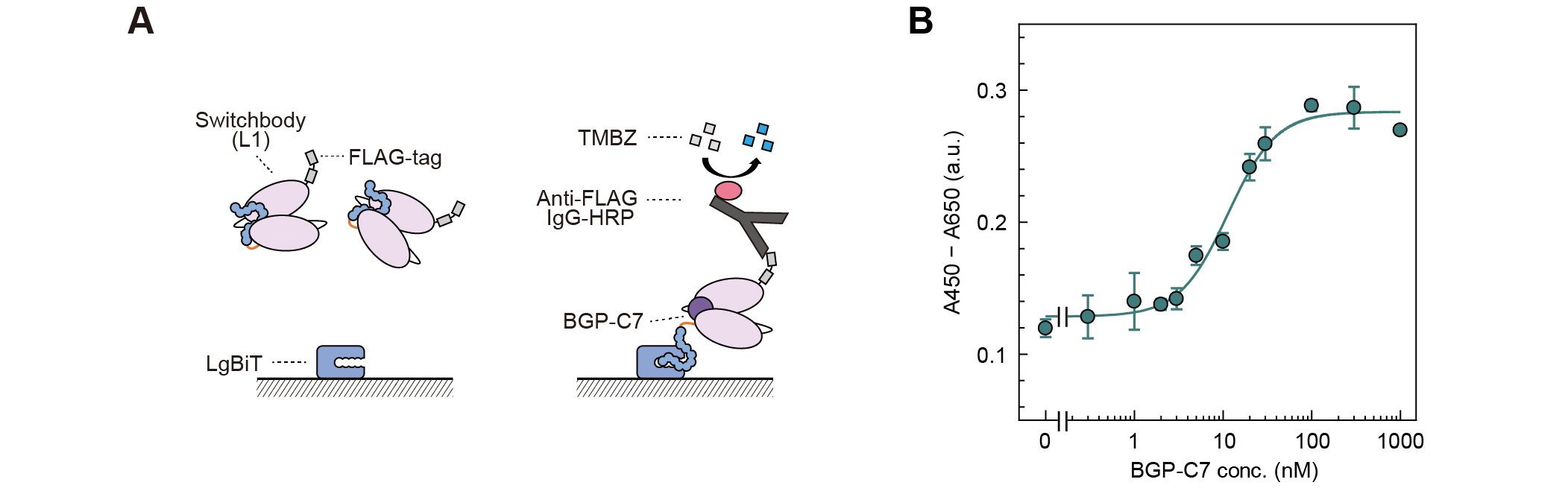
抗原添加による発光応答がHiBiTのトラップとリリースに基づくものかを調べるため、固定化したLgBiTに対するSwitchbodyの結合をELISAで評価したところ、BGP-C7を添加すると濃度に応じてシグナルが増加した(図2A, B)。これは、抗原非存在下で抗原結合部位近傍にトラップされていたHiBiTが、抗原結合によってLgBiTとより相互作用しやすい状態となったことを示しており、我々が想定した作動原理「トラップ&リリース」モデルを支持している。
 |
|
| 図2. | (A)LgBiTを固定化したELISAの模式図(B)抗原濃度依存的なシグナルの上昇(DOI:10.1002/advs.202508422より一部改変) |
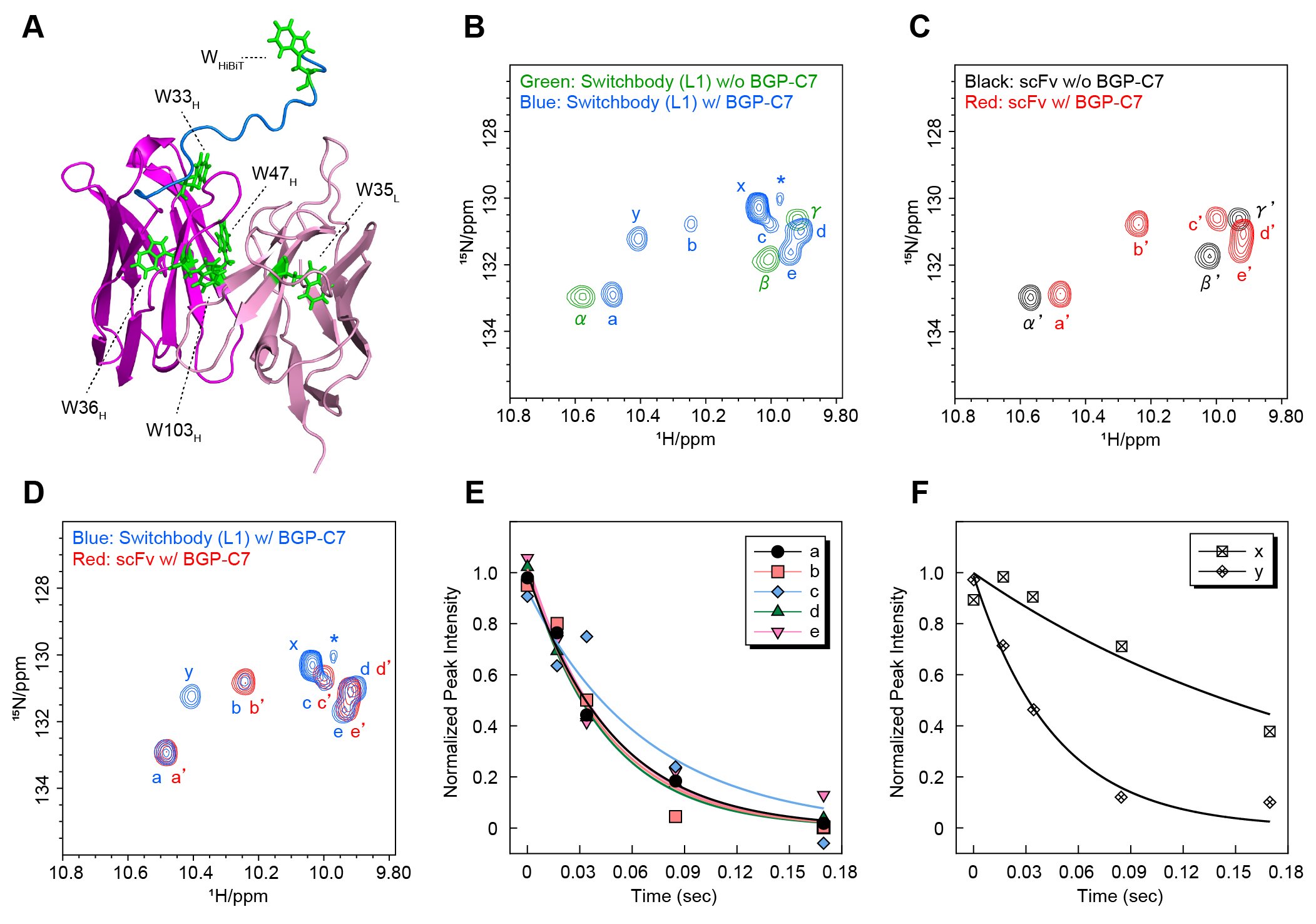
さらに、詳細に作動原理を調べるため、溶液中におけるタンパク質の構造変化や運動性に関する情報を得られるNMR解析を行った。HiBiTにはトリプトファン残基が1つ含まれているため(図3A)、1H-15N HSQCスペクトルを測定し、一般にトリプトファン側鎖のNHに由来するシグナルが現れるとされる領域に着目して解析を行った。SwitchbodyとHiBiTをもたないscFvについて、抗原非存在下では限られた数のシグナルのみが観測されたのに対し、抗原を添加すると新たなシグナルが現れた(図3B青, C赤)。さらに、抗原存在下のSwitchbodyとscFvのスペクトルを比較したところ、Switchbodyに特有のシグナルx、yが確認された(図3D)。続いて横緩和時間T2を測定し、各シグナルの運動性を評価した。シグナルxは他のシグナルと比べて横緩和速度が小さく、より高い運動性をもつことが示された(図3E, F)。このことから、シグナルxは抗原結合によって運動性が増加したHiBiT中のトリプトファンに由来すると考えられた。一方、シグナルyの横緩和速度はscFv由来と考えられるシグナルa-eと近い値を示したため、HiBiTそのものではなく、HiBiTとの相互作用によって影響を受けたscFv領域のトリプトファンに由来する可能性が高いと考えられた。これらの結果から、scFvのN末端側に融合されたHiBiTは、抗原非存在下では抗原結合部位近傍で運動性が抑えられている一方、抗原結合によってより運動性が高い状態へ移行することが強く示唆された。当初想定したとおり、抗原非存在下でHiBiTが抗原結合部位近傍にトラップされ、抗原結合によってリリースされることが、NMR解析の結果からも支持された。
 |
|
| 図3. | (A)結晶構造(PDB ID:5X5X)をもとに作製したSwitchbodyのモデル構造(B-D)1H-15N HSQCスペクトルの重ね合わせ(E, F)抗原存在下での横緩和時間T2の測定結果(DOI:10.1002/advs.202508422より転載) |
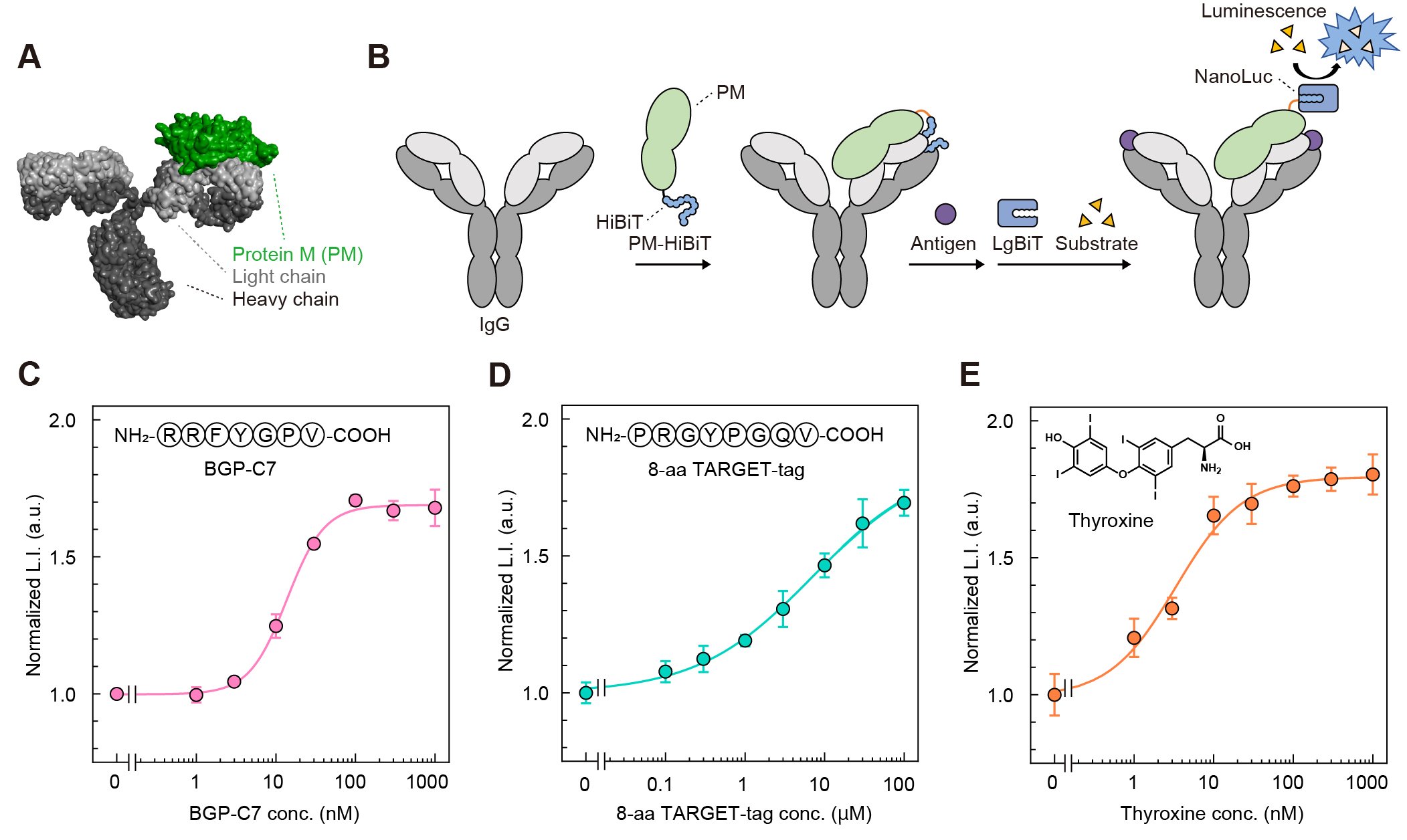
最後に、「トラップ&リリース」に基づく酵素活性の制御が、KTM219以外の抗体にも応用できるかを検討した。多様な抗体を評価するうえで、抗体ごとに遺伝子改変を行ってHiBiTを融合する方法は、多くの手間と時間を要する。そこで、抗体結合タンパク質Protein M(PM)にHiBiTを融合したプローブPM-HiBiTを作製した(図4A, B)。Protein Mは多くのIgG抗体の軽鎖に結合できるため、このプローブを用いれば、市販のIgG抗体を遺伝子改変せずに発光型Switchbodyへ変換できることを期待した。PM-HiBiTを、KTM219のIgG、ペプチドタグであるTARGET-tagを認識するIgG、甲状腺ホルモンであるチロキシンを認識するIgGとそれぞれ混合し、抗原を添加したところ、いずれの抗体を用いた場合でも、発光強度が抗原濃度に応じて増加した(図4C-E)。これにより、HiBiTの「トラップ&リリース」による発光応答は特定の抗体に限定されず、多様な抗体へ展開できる可能性が示された。さらに、SwitchbodyはNanoLucに限らず、他の酵素へも拡張できる可能性がある。実際に我々は、酵素を変更したSwitchbodyの開発を進めており、未発表データではあるが、異なる酵素ですでに抗原依存性酵素スイッチとして機能することも確認している。
 |
|
| 図4. | (A)結晶構造(PDB ID:4NZR, 1IGT)をもとに作製したPM/抗体複合体のモデル構造(B)PMに融合されたHiBiTの抗体によるトラップと抗原結合によるリリースの模式図(C-E)PM-HiBiT/抗体複合体に抗原を添加したときの発光強度変化(DOI:10.1002/advs.202508422より転載) |
タンパク質をエンジニアリングして新たな機能を持たせる技術は、構造予測や生成AIの進展によって、経験的な試行錯誤から予測に基づく設計へと移りつつある。抗体の分子認識や酵素機能も、今後さらに精密に設計できるようになると期待される。Switchbodyは、抗体が「何を認識するか」と、酵素が「何を起こすか」を結びつける分子ツールである。今後、抗体と酵素の設計がさらに発展すれば、研究者が選んだ分子を認識し、必要な機能を作動させるタンパク質スイッチを、より速く、より自在に作り出せるようになるかもしれない。
|
参考文献 |
|
| [1] | T. Yasuda, Y. Ueno, M. Taguchi, N. Tochio, H. Yagi, S. Yazaki, R. Arai, B. Zhu, T. Kigawa, H. Ueda, T. Kitaguchi Adv. Sci. 12, e08422 (2025) (DOI: 10.1002/advs.202508422) |
| 関連リンク | |
| ●Science Tokyo News:https://www.isct.ac.jp/ja/news/ospry020twom | |
| ●北口研究室:https://kitaguchi.jimdofree.com/ |
