
Latest Research
- 2026.06.29
- Kitaguchi Group
Antibody Engineering from Measurement to Manipulation
Biological phenomena are orchestrated by the functions of a wide variety of proteins. For example, membrane receptor proteins on the cells recognize extracellular stimuli and changes in the environment, and transmit as intracellular signals. The cell signaling regulates diverse cellular functions, including cell proliferation, differentiation, metabolism, cell death, and exocytosis, and serves as an essential mechanism underlying complex biological phenomena such as development, morphogenesis, homeostasis, tissue repair, and neuronal activity. Therefore, manipulating signaling pathways will make it possible to investigate how they regulate diverse cellular functions, thereby deepening our understanding of biological phenomena. Furthermore, if cellular functions impaired by disrupted signaling pathways can be appropriately restored, this may also lead to the novel therapeutic strategies.
In recent years, the chemogenetic tool, DREADD has become widely used. It manipulates cellular functions through artificially engineered receptors with small-molecule ligands that selectively activate those receptors. Although this tool enables external control of specific signaling pathways, the variation of ligands is limited. Therefore, there is a need for tools that enable scientists to manipulate cellular functions using ligands of their choice. To achieve this, a new strategy was required for designing protein switches that recognize and respond to various molecules. In this study, we developed an enzyme switch, Switchbody, which exploits the high molecular recognition ability of antibodies to regulate enzyme activity in response to antigen binding.[1] Because enzymes regulate various intracellular signaling pathways, extending this design strategy to enzymes responsible for the synthesis or degradation of intracellular signaling molecules will enable new tools for controlling cellular functions in response to the recognition of molecules of interest.
The design strategy for Switchbody was inspired by the working principle of the fluorescent immunosensor, Q-body. In Q-bodies, a fluorescent dye labeled to a N-terminus of antibody is trapped near the antigen-binding site, resulting in fluorescence quenching. Upon antigen binding, the fluorescent dye is released, causing an increase in fluorescence intensity, thereby enabling quantitative detection of antigen concentration through the fluorescence response. This working principle suggests that antibodies not only recognize antigens but also switch the state of a molecule positioned near the antigen-binding site in response to antigen binding. In this study, we attempted to extend this working principle from a fluorescent dye to an enzyme fragment by establishing a principle for translating antigen binding into changes in enzyme activity.
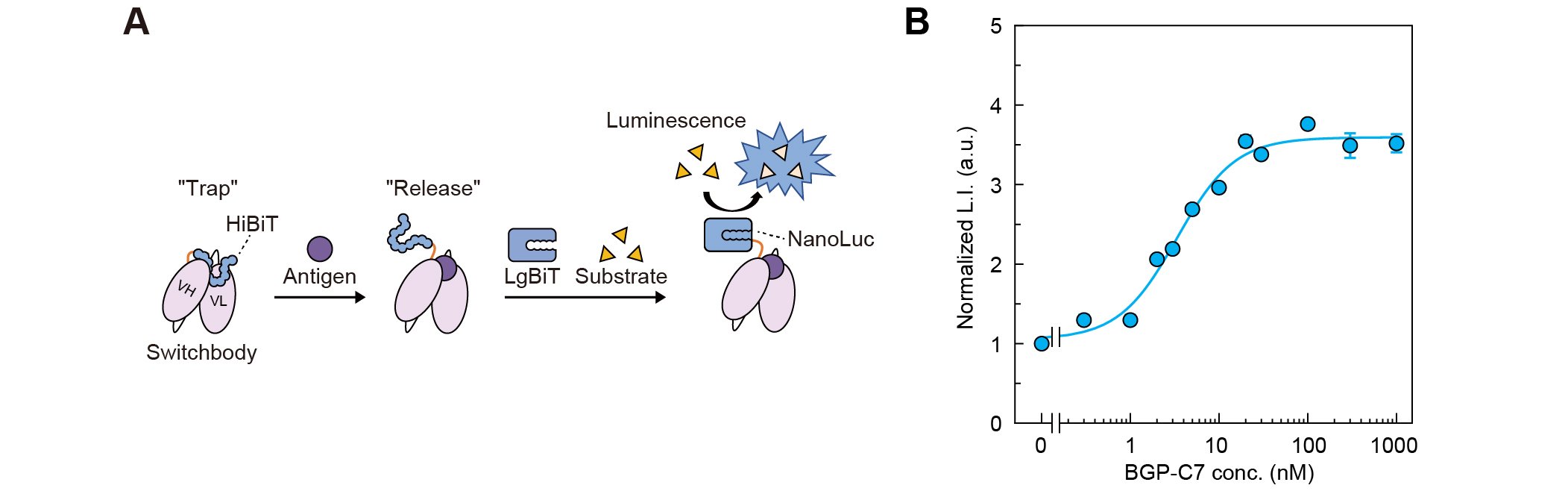
As a bioluminescent Switchbody, we designed a protein in which HiBiT, an 11-amino-acid peptide derived from the bioluminescent enzyme NanoLuc, was fused to the N-terminus of a scFv derived from KTM219, an antibody that recognizes the peptide antigen BGP-C7. In this design, we anticipated that HiBiT would be trapped near the antigen-binding site in the absence of antigen, preventing interaction with LgBiT, the 18-kDa subunit of NanoLuc. Upon antigen binding, HiBiT would be released and interact with LgBiT, reconstituting an enzyme and an increase in its activity (Fig. 1A). The HiBiT-fused scFv was expressed in Escherichia coli and purified. When luminescence intensity was measured in the presence of LgBiT and its substrate furimazine, the intensity increased in a dose-dependent manner in response to antigen, demonstrating that the HiBiT-fused scFv functioned as a bioluminescent Switchbody (Fig. 1B).
 |
|
| Figure 1. | (A) Schematic illustration of antigen detection by Switchbody. (B) Dose-dependent luminescence intensity increase of Switchbody. (Adapted with permission from DOI: 10.1002/advs.202508422) |
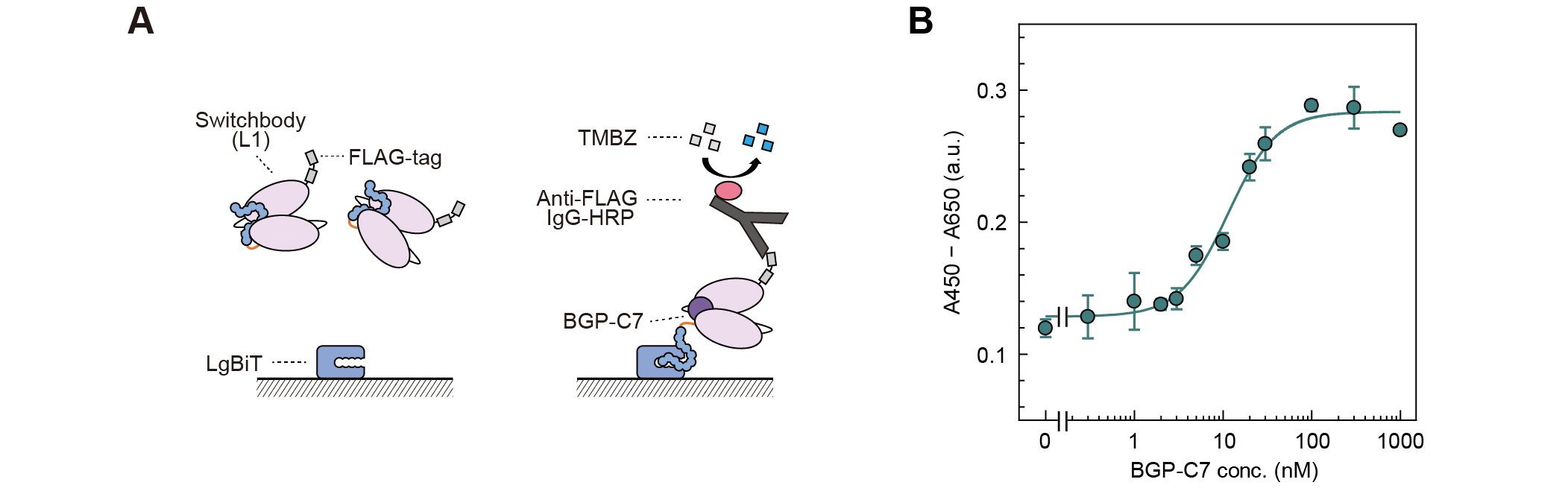
To investigate whether the antigen-induced luminescence response is based on the trap and release of HiBiT, we evaluated the binding of Switchbody to immobilized LgBiT by ELISA (Fig. 2A). Upon addition of BGP-C7, the absorbance signal increased in a dose-dependent manner (Fig. 2B). This result indicates that HiBiT, which is trapped near the antigen-binding site in the absence of antigen, becomes more accessible to interaction with LgBiT upon antigen binding, thereby supporting our proposed "Trap and Release" principle.
 |
|
| Figure 2. | (A) Schematic illustration of ELISA with immobilized LgBiT. (B) Dose-dependent absorbance signal increase. (Adapted with permission from DOI: 10.1002/advs.202508422) |
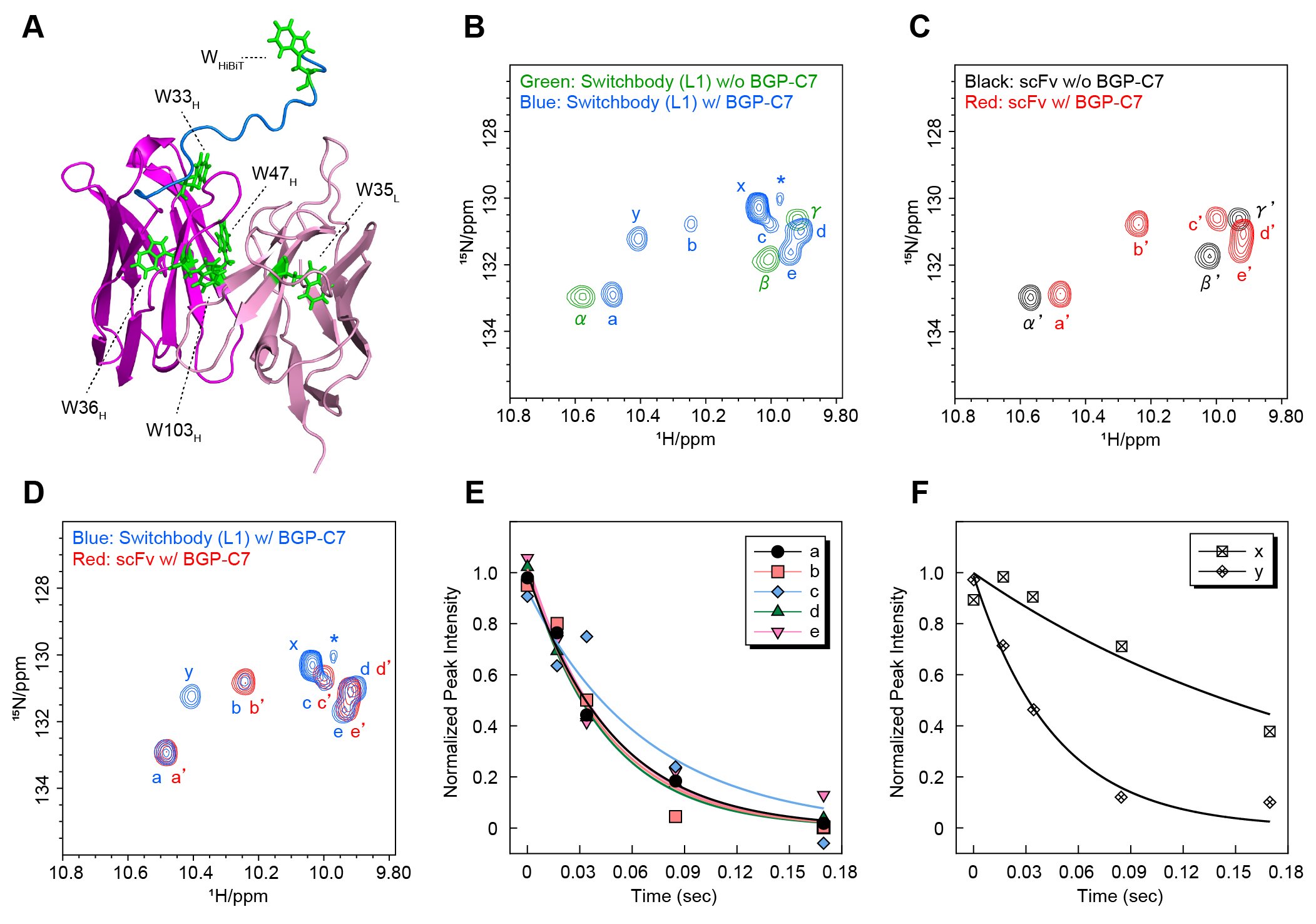
To further investigate the working principle in detail, we performed nuclear magnetic resonance (NMR) analyses, which provide information on protein structural changes in solution. Since HiBiT contains a single tryptophan residue (Fig. 3A), we measured 1H-15N HSQC spectra and focused on the region where signals derived from the NH group of tryptophan side chains are generally observed. For both Switchbody and scFv, three signals were observed in the absence of antigen, whereas additional signals appeared upon antigen addition (Fig. 3B, blue; Fig. 3C, red). Furthermore, comparison of the spectra of Switchbody and scFv in the presence of antigen revealed two signals, designated x and y, that were unique to Switchbody (Fig. 3D). Next, to evaluate the dynamics of the HiBiT region of the Switchbody, we measured the transverse relaxation time (T₂) in the presence of antigen. Signal x exhibited a lower transverse relaxation rate than the other signals, indicating higher molecular mobility (Fig. 3E, F). This result is consistent with signal x originating from the tryptophan residue in HiBiT, and reflecting increased mobility upon antigen binding. In contrast, the transverse relaxation rate of signal y was comparable to those of signals a-e, which are presumed to be derived from the scFv. Therefore, signal y would arise from a tryptophan residue in the scFv, with its local environment affected through interaction with HiBiT. Taken together, these results suggest that HiBiT fused to the N-terminus of the scFv has restricted mobility near the antigen-binding site in the absence of antigen, whereas antigen binding induces a transition of HiBiT to a more mobile state. The NMR analyses support our hypothesis that HiBiT is trapped near the antigen-binding site in the absence of antigen and released upon antigen binding.
 |
|
| Figure 3. | (A) Model 3D structure of Switchbody generated from the crystal structure (PDB ID: 5X5X). (B-D) Overlays 1H-15N HSQC spectra. (E, F) Results of T2 measurements in the presence of antigen. (Reproduced with permission from DOI: 10.1002/advs.202508422) |
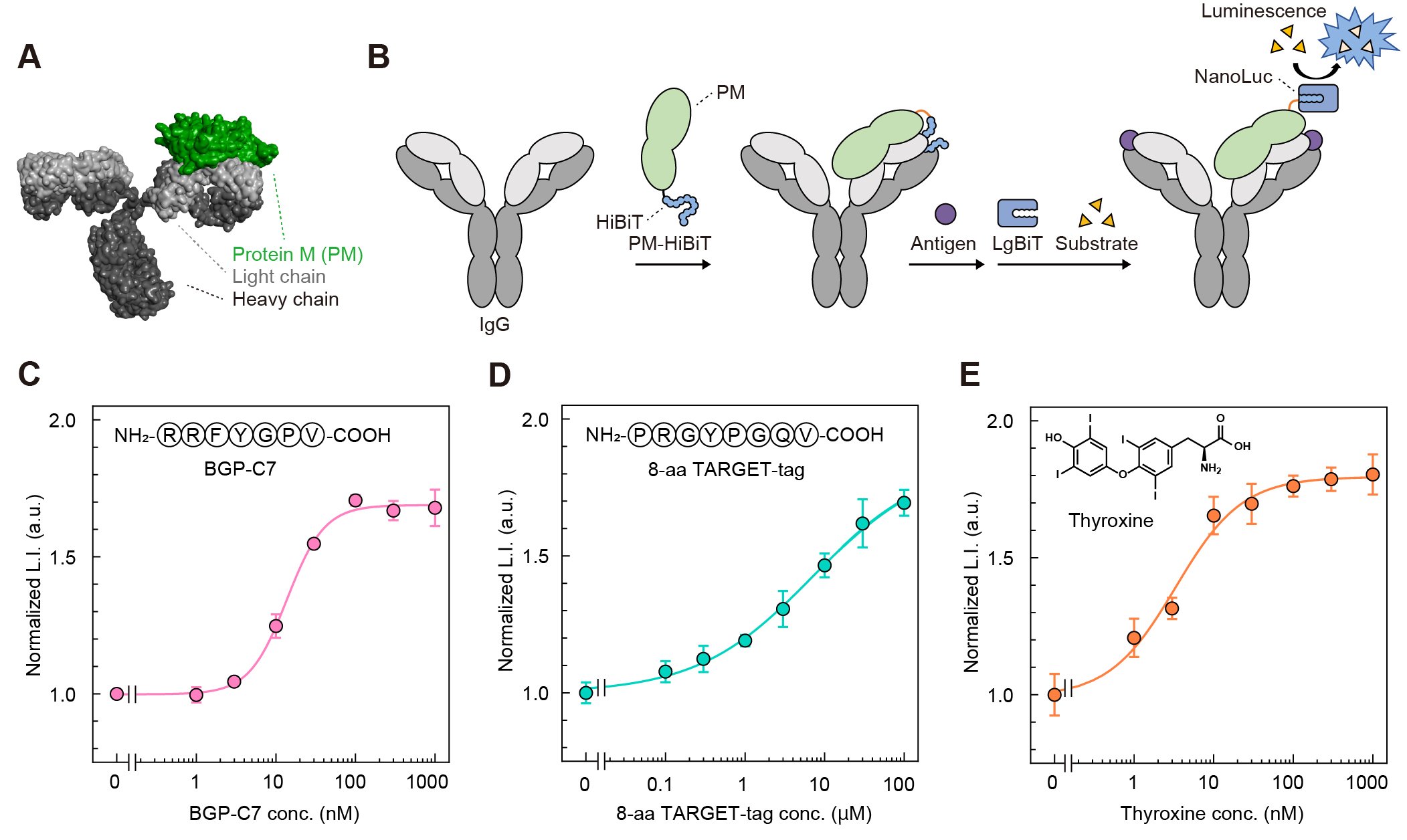
Finally, we examined whether regulation of enzyme activity based on the "Trap and Release" of enzyme fragments is applicable to antibodies other than KTM219. Evaluating various antibodies by genetically manipulating each antibody to fuse HiBiT requires considerable time and effort. Therefore, we developed PM-HiBiT, in which HiBiT was fused to the antibody-binding protein Protein M (PM) (Fig. 4A, B). Since PM binds to the light chains of a wide variety of IgGs, we expected that PM-HiBiT enable commercially available IgGs to be converted into bioluminescent Switchbodies without genetic manipulation. PM-HiBiT was mixed with an IgG of KTM219, an IgG recognizing the TARGET-tag, or an IgG recognizing the thyroid hormone thyroxine, followed by addition of the corresponding antigen. In all cases, the luminescence intensity increased in a dose-dependent manner (Fig. 4C-E), suggesting the versatility of "Trap and Release" principle of HiBiT. Furthermore, the enzyme employed for Switchbody is not limited to NanoLuc. We are currently developing Switchbodies based on other enzymes and have confirmed that Switchbodies fusing other enzymes also function as antigen-responsive enzyme switches.
 |
|
| Figure 4. | (A) Model 3D structure of PM/IgG complex generated from their crystal structures (PDB ID: 4NZR and 1IGT). (B) Schematic illustration of trap of HiBiT fused to PM and its release upon antigen binding. (C-E) Luminescence responses of PM-HiBiT/IgG complexes upon antigen addition. (Reproduced with permission from DOI: 10.1002/advs.202508422) |
Technologies for engineering proteins to acquire new functions are shifting from empirical trial-and-error approaches to prediction-based design, driven by advances in structure prediction and generative artificial intelligence. Molecular recognition by antibodies and enzymatic functions are also expected to become more precisely designable in the future. Switchbody is a tool that links "what an antibody recognizes" with "what an enzyme does". As technology of antibody and enzyme design continue to advance, it may become possible to create protein switches more rapidly and more flexibly that recognize molecules selected by scientists and activate desired functions.
|
Reference |
|
| [1] | T. Yasuda, Y. Ueno, M. Taguchi, N. Tochio, H. Yagi, S. Yazaki, R. Arai, B. Zhu, T. Kigawa, H. Ueda, T. Kitaguchi Adv. Sci. 12, e08422 (2025) (DOI: 10.1002/advs.202508422) |
| Related links | |
| ●Science Tokyo News: https://www.isct.ac.jp/en/news/yuehggovajdb | |
| ●Kitaguchi Group: https://kitaguchi.jimdofree.com/ |
